

COMPONENTES CELULARES DO SISTEMA NERVOSO
O sistema nervoso é o responsável pela nossa capacidade de aprender, sentir, lembrar e nos movimentar. Todo pensamento, ação e emoção refletem a atividade do sistema nervoso. Porém, a atividade do sistema nervoso também auxilia na manutenção das condições do meio interno do nosso corpo. Assim, o sistema nervoso também constitui um dos principais sistemas de controle responsáveis pela manutenção da homeostasia corporal.
O sistema nervoso é único e altamente integrado. Porém, por conveniência didática, ele é dividido em duas partes: o sistema nervoso central (SNC) e o sistema nervoso periférico (SNP). O SNC compreende o encéfalo e a medula espinal. Ele ocupa a cavidade dorsal do corpo, estando localizado dentro da cabeça (encéfalo) e da coluna vertebral (medula espinal). O SNP, por sua vez, compreende toda a parte do sistema nervoso fora do SNC. Ele é composto pelos nervos (cranianos e espinais) e seus gânglios (Figura 1).

Figura 1: Divisões do sistema nervoso. Observe, à direita, o encéfalo e a medula espinal, que compõem o sistema nervoso central (SNC). À esquerda, estão os componentes do sistema nervoso periférico (SNP), que são os nervos (cranianos e espinais) e seus gânglios (Fonte: modificado de https://commons.wikimedia.org/wiki/File:1201_Overview_of_Nervous_System_esp.jpg).
O sistema nervoso é uma rede de bilhões ou trilhões de células nervosas ligadas umas às outras de forma extremamente organizada, de modo a ter um rápido controle sobre os sistemas do corpo. Os tipos celulares principais do sistema nervoso são as células nervosas ou neurônios e as células da glia ou células neurogliais. As células da glia são diferentes no sistema nervoso central (astrócitos, oligodendrócitos, micróglia, células ependimárias) e no sistema nervoso periférico (células de Schwann nos nervos periféricos e as células-satélite nos gânglios).
As palavras neurônio e glia têm origem no grego, a partir das palavras gregas neûron, que significa “nervo”, e glí(ā), que significa “cola”. O sistema nervoso humano possui aproximadamente 86 bilhões de neurônios, sendo o número de células da glia igual ou maior do que o de neurônios. Os neurônios são as células especializadas na sinalização elétrica por longas distâncias (geram e conduzem potenciais de ação). As células da glia se comunicam com os neurônios e fornecem a estes um importante suporte físico e bioquímico. Portanto, os neurônios e as células da glia são essenciais na comunicação interna do sistema nervoso.
As células da glia presentes no SNC são a micróglia ou microglia, os oligodendrócitos, os astrócitos e as células ependimárias (Figura 2).

Figura 2: Células da glia localizadas no sistema nervoso central. No esquema são mostrados astrócitos com prolongamentos envolvendo um capilar sanguíneo, oligodendrócitos formando internodos de bainha de mielina em torno de axônios, micróglia e células ependimárias (Fonte: Modificado de https://commons.wikimedia.org/wiki/File:Glial_Cell_Types.png).
Os astrócitos (Figura 2) constituem o maior número de células gliais do SNC. Seu nome deriva de seus corpos celulares irregulares, semelhantes a estrelas. Além disso, o grande número de prolongamentos dos astrócitos os colocam em contato tanto com neurônios como com capilares sanguíneos. Estima-se que os astrócitos constituem cerca de metade das células do encéfalo, sendo encontrados por todo o SNC.
Os prolongamentos de astrócitos situados próximo da membrana de neurônios atuam abastecendo os neurônios com substratos para a produção de ATP, além de liberarem mediadores químicos como os fatores neurotróficos. Os prolongamentos de astrócitos próximos das sinapses desempenham funções principalmente na captura de íons potássio, de água e dos neurotransmissores glutamato e ácido γ-aminobutírico (GABA). Os astrócitos também possuem extremidades que cercam os vasos sanguíneos e fazem parte da barreira hematencefálica. Essa barreira impede a difusão de moléculas grandes ou hidrofílicas (por exemplo, proteínas) para o líquido cerebrospinal e para o parênquima do SNC, mas permite a passagem de moléculas pequenas e moléculas lipossolúveis. A barreira hematencefálica é constituída pelas junções oclusivas do endotélio capilar e pelos pés dos astrócitos que envolvem os capilares sanguíneos. Assim, os astrócitos desempenham importante papel na manutenção da homeostasia do líquido extracelular do SNC.
Você sabia?
Os astrócitos respondem à lesão traumática encefálica pelo aumento em seu tamanho e, em alguns casos, aumento também de sua quantidade. O processo é chamado astrocitose reativa e se caracteriza por aumento de células expressando a proteína ácida glial fibrilar (GFAP), pela síntese e liberação de citocinas que regulam respostas inflamatórias e a entrada de células hematogênicas no SNC.
A microglia (Figura 2) não se relaciona fisiológica e embriologicamente com os outros tipos de células da glia e com os neurônios, pois são células do sistema imune especializadas que residem permanentemente no SNC. Elas são originadas de macrófagos externos ao SNC, sendo, portanto, “células de limpeza” (scavenger) que se assemelham aos macrófagos de outros tecidos. Quando ativada, a microglia remove células danificadas e invasores, desempenhando papel importante em situações de lesão, infecção e doenças do SNC. Porém, essas células podem ser ativadas em outras situações e, como liberam diversos fatores neurotróficos quando ativadas, as células microgliais contribuem para o desenvolvimento da dor e de doenças neurodegenerativas.
As células ependimárias (Figura 2) formam uma camada epitelial com permeabilidade seletiva, o chamado epêndima, o qual separa os compartimentos líquidos do SNC. O epêndima é também uma fonte de células-tronco neurais, as quais são células imaturas que podem diferenciar-se em neurônios e em células da glia.
Os oligodendrócitos (Figuras 2, 4 e 5) são as células responsáveis pela formação da bainha de mielina que envolve os axônios no SNC. A mielina é uma substância composta principalmente por várias camadas concêntricas de fosfolipídeos de membrana localizadas em torno dos axônios. Sendo composta predominantemente por componentes lipídicos, a mielina não conduz sinais elétricos, por isso atua como isolante elétrico. Ao longo de um axônio formam-se regiões com mielina alternadas com espaços sem mielina (Figuras 4, 5 e 7). As lacunas localizadas entre as porções de bainhas de mielina vizinhas recebem o nome de nós de Ranvier. Nesses nós, portanto, uma pequena porção do axônio é exposta ao espaço intersticial o que permite a troca de íons entre o líquido intracelular (LIC) e o líquido extracelular (LEC), condição necessária para a geração de potenciais de membrana. Nos nós de Ranvier há acúmulo de canais de sódio regulados por voltagem. Assim, entre os nós de Ranvier, a mielina isola eletricamente o axônio do espaço extracelular enquanto nos nós de Ranvier não há isolamento e podem ocorrer potenciais de ação. Além disso, pode haver a disseminação eficiente da despolarização entre um nó e o outro. Isso permite que os potenciais de ação se propaguem rapidamente, saltando de um nó para o próximo, um processo chamado de condução saltatória. Revise o capítulo sobre a geração e propagação dos potenciais de membrana.
As células da glia encontradas no SNP são as células-satélite e as células de Schwann (Figuras 3, 4 e 5). As células-satélite são as células que formam cápsulas de suporte ao redor dos corpos celulares dos neurônios localizados em gânglios (Figura 3), enquanto as células de Schwann são as responsáveis pela formação da bainha de mielina que envolve os axônios no SNP (Figuras 4 e 5).

Figura 3: Células da glia localizadas no sistema nervoso periférico. Observe as células de Schwann formando a mielina em torno do axônio e as células satélites no gânglio (Fonte: Modificado de https://commons.wikimedia.org/wiki/File:1210_Glial_Cells_of_the_PNS.jpg).
Uma diferença entre células de Schwann e oligodendrócitos é o número de axônios que cada célula envolve (Figuras 4 e 5). No SNC, um oligodendrócito se ramifica e forma mielina em volta de uma porção de axônios de diferentes neurônios. Uma célula de Schwann, por sua vez, se associa a apenas uma determinada porção de um único axônio. Assim, um único axônio no SNP pode possuir mais de 500 células de Schwann ao longo do seu comprimento, pois cada célula de Schwann envolve um segmento de cerca de 1 a 1,5 mm. Do mesmo modo que no SNC, os nós de Ranvier no SNP são áreas localizadas entre os segmentos isolados eletricamente pela presença da mielina (Figura 5).

Figura 4: Formação da camada de mielina em axônios do sistema nervoso central (SNC) e do sistema nervoso periférico (SNP). O oligodendrócito (SNC) e as células de Schwann (SNP) formam internodos de mielina em torno do axônio. Porém, o oligodendrócito se ramifica e forma internodos de mielina ao redor de uma porção de diferentes axônios, enquanto cada célula de Schwann forma um dos internodos de um único axônio (Fonte: Modificado de https://upload.wikimedia.org/wikipedia/commons/5/52/CeluladeSchwann.jpg; https://commons.wikimedia.org/wiki/File:Oligodendrocyte_illustration.png).
O neurônio é um tipo celular diferenciado para realizar a comunicação intercelular. Esta célula constitui a unidade funcional do sistema nervoso e é especializada em gerar sinais elétricos e conduzir mensagens na forma de impulsos nervosos (potenciais de ação). Cada neurônio possui o corpo celular (ou soma) com núcleo que contém o material genético e citoplasma com organelas necessárias para a atividade celular. Do corpo celular se estendem longos prolongamentos que, de modo geral, são dendritos e axônio (Figura 5 e 7).

Para pensar e pesquisar: Como a informação é codificada no sistema nervoso? Sabemos que são usados sinais elétricos e sinais químicos. Os sinais elétricos são os chamados potenciais de membrana e os neurotransmissores atuam como sinais químicos. Em que parte do neurônio se formam os potenciais de ação? E os sublimiares? Como eles se relacionam? Leia os capítulos sobre bioeletrogênese e potenciais de membrana para revisar como se formam esses potenciais, suas características e funções.

Figura 5: Partes do neurônio e formação da camada de mielina em axônios do sistema nervoso central (SNC) e do sistema nervoso periférico (SNP). O oligodendrócito (SNC) e as células de Schwann (SNP) formam internodos de mielina em torno do axônio. Porém, o oligodendrócito se ramifica e forma internodos de mielina ao redor de uma porção de diferentes axônios, enquanto cada célula de Schwann forma um dos internodos de um único axônio.
Os dendritos são prolongamentos finos e ramificados, os quais são os alvos primários de sinais de entrada que chegam de outros neurônios. Os dendritos, portanto, aumentam a área de superfície receptora do neurônio ao possibilitar que este se comunique com muitos outros neurônios. Muitos dendritos possuem ainda os chamados espinhos dendríticos, que podem variar de espinhos finos até botões com formato de cogumelo. Quando presentes, os espinhos dendríticos expandem ainda mais a área de recepção de estímulos do neurônio (Figura 6). Os espinhos dendríticos podem alterar seu tamanho e formato em resposta a um sinal de entrada de células vizinhas.

Figura 6: Espinhos dendríticos. Observe à esquerda um dendrito de um neurônio do estriado (SNC) com alguns espinhos. Dendrite: dendrito. Spine neck: colo ou pescoço do espinho. Spine head: cabeça do espinho. Para dimensionar o tamanho dos espinhos veja a barra de escala (acima, à direita) indicando 1 micrômetro. À direita, podem ser vistos dendritos e espinhos dendríticos de um neurônio do hipocampo (SNC) em cultura celular, que expressa GFP (proteína fluorescente verde). (Fonte: https://commons.wikimedia.org/wiki/Category:Dendritic_spines#/media/File:Dendritic_spines.jpg/2; https://commons.wikimedia.org/wiki/Category:Dendritic_spines#/media/File:Dendriticspines.jpg).
Você sabia?
As alterações na morfologia dos espinhos dendríticos são associadas tanto a processos de aprendizagem e memória quanto a várias patologias, incluindo alterações genéticas que ocasionam deficiência intelectual e doenças neurodegenerativas, como a doença de Alzheimer.
O axônio (Figura 7) é um prolongamento único especializado em transmitir sinais do neurônio para outros neurônios ou do neurônio para células musculares e endócrinas. O axônio emerge do corpo do neurônio em uma região especializada denominada cone axonal ou cone de implantação. Logo após emergir, o axônio se estreita formando um prolongamento delgado que mantém seu diâmetro uniforme pelo resto de seu comprimento. Porém, o axônio pode ocasionalmente se ramificar ao longo de seu comprimento. As ramificações são chamadas colaterais axonais, os quais emergem do axônio tronco em ângulos mais ou menos retos. Tendo ou não esses colaterais, um axônio geralmente se ramifica profusamente em sua região terminal (Figura 7). As regiões terminais dilatadas das ramificações terminais recebem o nome de terminais axônicos, protuberâncias ou botões sinápticos.
Figura 7: Estrutura de um neurônio, mostrando corpo celular, núcleo, dendritos e axônio, com seu cone de implantação, segmento inicial e terminal axonal. Pode-se observar neste axônio a bainha de mielina e os nós de Ranvier (Fonte: Aprendendo Fisiologia).
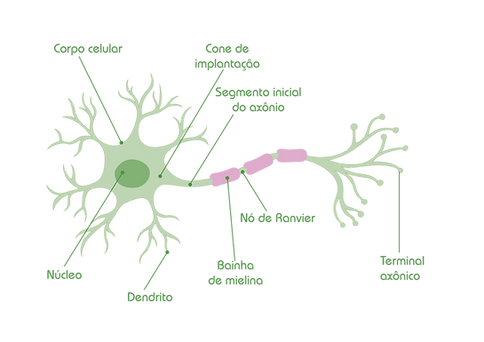
Os axônios exibem comprimentos que variam de poucas centenas de micrômetro (um micrômetro é um milionésimo de metro) até muito além, podendo chegar até a 1 metro de comprimento, dependendo do tipo de neurônio e do tamanho da espécie onde ele se encontra. Os axônios relativamente curtos são uma característica de neurônios de circuito local ou interneurônios. Os axônios de neurônios de projeção estendem-se para alvos distantes e seu comprimento é variável. Como informado previamente, os axônios podem possuir ou não bainha de mielina. O axônio com bainha de mielina é chamado axônio mielinizado, enquanto o axônio sem bainha de mielina recebe o nome de axônio amielínico ou não mielinizado. Os locais onde ocorrem axônios mielinizados com poucos corpos celulares recebe a denominação de substância branca. Na substância cinzenta encontram-se os corpos celulares dos neurônios, os dendritos e os terminais axônicos.
Diferentemente do corpo neuronal e dos dendritos, o axônio não possui corpúsculos de Nissl e aparelho de Golgi, estruturas envolvidas com a síntese e o empacotamento de proteínas (Figuras 5 e 7). Consequentemente, um axônio depende de seu corpo celular para sintetizar as proteínas necessárias para seu funcionamento e os componentes das membranas. Por este motivo, os axônios sofrem um rápido declínio se forem cortados ou severamente danificados. Além disso, é preciso um mecanismo eficiente de transporte para distribuir essas proteínas.
Todas as proteínas do terminal axônico são sintetizadas no corpo celular e chegam ao terminal por transporte axonal. O transporte axonal lento transporta componentes que não são consumidos rapidamente pela célula, como as enzimas e as proteínas do citoesqueleto celular, em velocidade de 0,2 a 2,5 mm/dia. A direção desse transporte é do corpo celular para o terminal axônico.
O transporte axonal rápido transporta organelas em velocidades de até 400 mm/dia. Esse tipo de transporte ocorre em duas direções: o transporte anterógrado (para frente) transporta vesículas e mitocôndrias do corpo celular para o terminal axônico, enquanto o transporte retrógrado (para trás) transporta componentes celulares velhos para reciclagem, portanto, do terminal axônico para o corpo celular. O transporte retrógrado é parte da fisiopatologia de muitas doenças, incluindo pólio, raiva, tétano e herpes simplex.
Você sabia?
O transporte axonal é importante para a regeneração de nervos após uma lesão no SNP. Nesse caso, a porção do nervo acoplada ao corpo celular se fechará em horas e botões ou brotos aparecerão em um ou dois dias. A extremidade se encherá com mitocôndrias e retículo endoplasmático liso. O brotamento crescerá como finas fibras. Se a regeneração for bem-sucedida, uma das novas fibras encontrará seu caminho em direção ao envoltório distal do nervo em degeneração e irá reinervar o alvo. Então, a fibra aumentará seu diâmetro e se tornará remielinizada.
Estruturalmente, os neurônios são classificados pelo número de prolongamentos originados no corpo celular (Figura 8). Os neurônios anaxônicos não possuem axônio identificável, mas possuem inúmeros dendritos ramificados. Os neurônios bipolares possuem apenas um axônio e um dendrito se estendendo a partir do corpo celular. Os neurônios pseudounipolares possuem o corpo celular localizado lateralmente com um único processo curto emergindo do mesmo, o qual logo se ramifica em forma de T em um prolongamento longo denominado axônio, pois durante o desenvolvimento o dendrito se fundiu e tornou parte do axônio. Um neurônio multipolar eferente típico possui de 5 a 7 dendritos, cada um se ramificando de 4 a 6 vezes, e um único axônio longo, que pode emitir colateral diversas vezes.

Figura 8: Classificação dos neurônios quanto ao número de prolongamentos que emergem do corpo celular (Fonte: modificado de https://commons.wikimedia.org/wiki/File:Neuron_Classification.png).
Outra classificação dos neurônios os agrupa de acordo com a direção que os impulsos nervosos trafegam em relação ao SNC (Figura 9). Com base nesse critério há neurônios sensoriais (aferentes), interneurônios e neurônios motores (eferentes). Os neurônios sensoriais (aferentes) transmitem impulsos de células receptoras sensoriais da pele e órgãos sensoriais internos em direção ou para dentro do SNC. Os neurônios motores transmitem impulsos para longe do SNC, para os órgãos efetores (músculos e glândulas) da periferia do corpo. Os interneurônios, também chamados neurônios de associação, localizam-se, em sua maioria, no SNC, geralmente entre os neurônios sensoriais e os neurônios motores de uma via neural e enviam sinais para vias do SNC envolvidas na integração de informações. Os interneurônios constituem 99% dos neurônios do corpo, incluindo aqueles do SNC. Quase todos são do tipo multipolar, porém com muitas ramificações complexas dos prolongamentos e sem longas extensões.

Figura 9: Classificação dos neurônios de acordo com a direção dos impulsos nervosos em relação ao sistema nervoso central. No esquema, são mostradas a via aferente (via que conecta um receptor sensorial a um centro integrador, no caso o sistema nervoso central) e a via eferente (sinal de saída que viaja do centro integrador, no caso o sistema nervoso central, para um efetor) (Fonte: Aprendendo Fisiologia).
Os neurônios não funcionam de forma isolada. Eles se organizam em conjuntos denominados circuitos neurais, os quais processam tipos específicos de informação e constituem as bases das sensações, percepções e comportamento. Os circuitos neurais podem variar de acordo com a sua função, mas normalmente são compostos por neurônios aferentes e eferentes e interneurônios. De modo geral, os circuitos que processam tipos semelhantes de informação compõem sistemas neurais que servem a propósitos comportamentais mais amplos. Geralmente, esses conjuntos são divididos em: 1) sistemas sensoriais, que adquirem e processam informação dos ambientes externo e interno de um indivíduo; 2) sistemas motores, que respondem às informações sensoriais gerando movimentos e outros comportamentos; e 3) sistemas associativos, que são responsáveis pelas funções encefálicas mais complexas.